There is something wonderful about time at sea, where your primary obligation is to observe the ocean from sunrise to sunset, day after day, scanning for signs of life. After hours of seemingly empty blue with only an occasional albatross gliding over the swells on broad wings, it is easy to question whether there is life in the expansive, blue, offshore desert. Splashes on the horizon catch your eye, and a group of dolphins rapidly approaches the ship in a flurry of activity. They play in the ship’s bow and wake, leaping out of the swells. Then, just as quickly as they came, they move on. Back to blue, for hours on end… until the next stirring on the horizon. A puff of exhaled air from a whale that first might seem like a whitecap or a smudge of sunscreen or salt spray on your sunglasses. It catches your eye again, and this time you see the dark body and distinctive dorsal fin of a humpback whale.
Figure 1. Pacific white-sided dolphins (Lagenorhynchus obliquidens) play in the big swell and surf the wake of the NOAA ship Bell M. Shimada off Coos Bay, Oregon. Photos: Dawn Barlow.
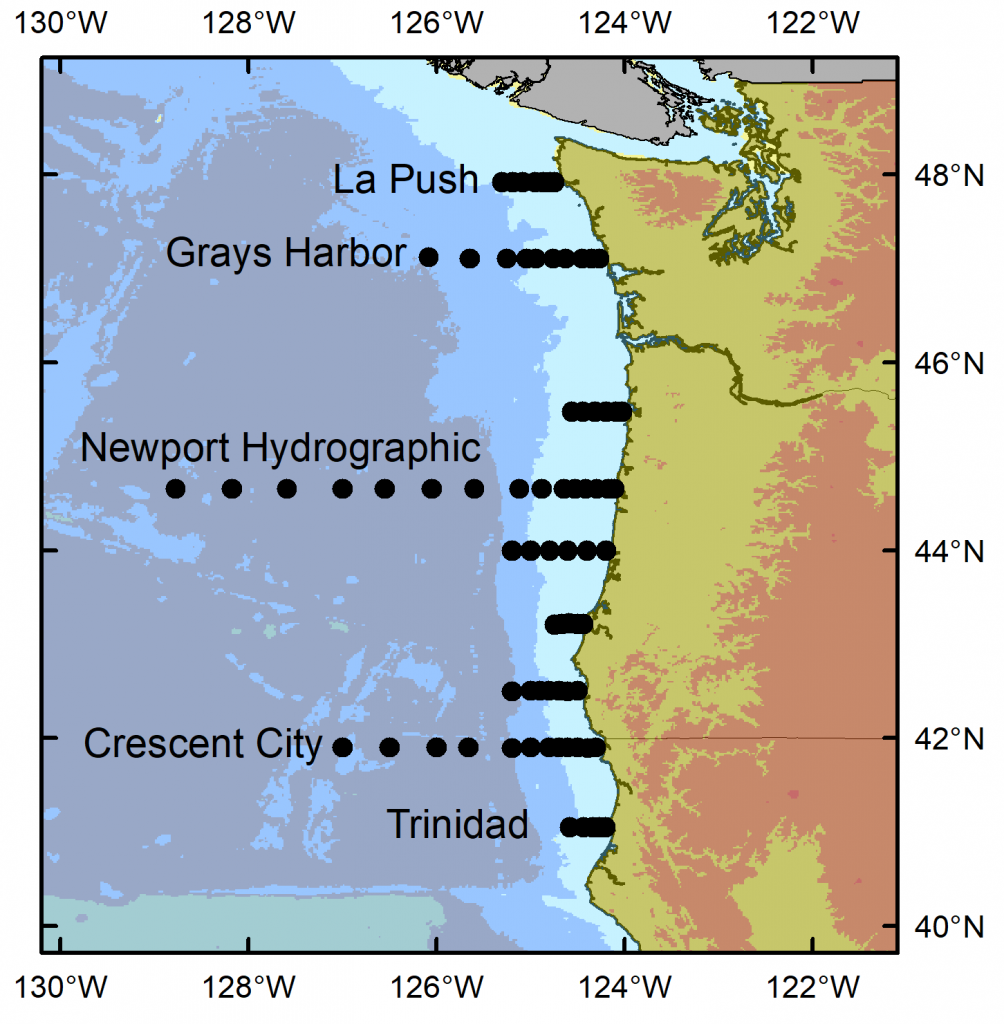
I have just returned from 10 days aboard the NOAA ship Bell M. Shimada, where I was the marine mammal observer on the Northern California Current (NCC) Cruise. These research cruises have sampled the NCC in the winter, spring, and fall for decades. As a result, a wealth of knowledge on the oceanography and plankton community in this dynamic ocean ecosystem has been assimilated by a dedicated team of scientists (find out more via the Newportal Blog). Members of the GEMM Lab have joined this research effort in the past two years, conducting marine mammal surveys during the transits between sampling stations (Fig. 2).
Figure 2. Northern California Current cruise sampling locations, where oceanography and plankton data are collected. Marine mammal surveys were conducted on the transits between stations.
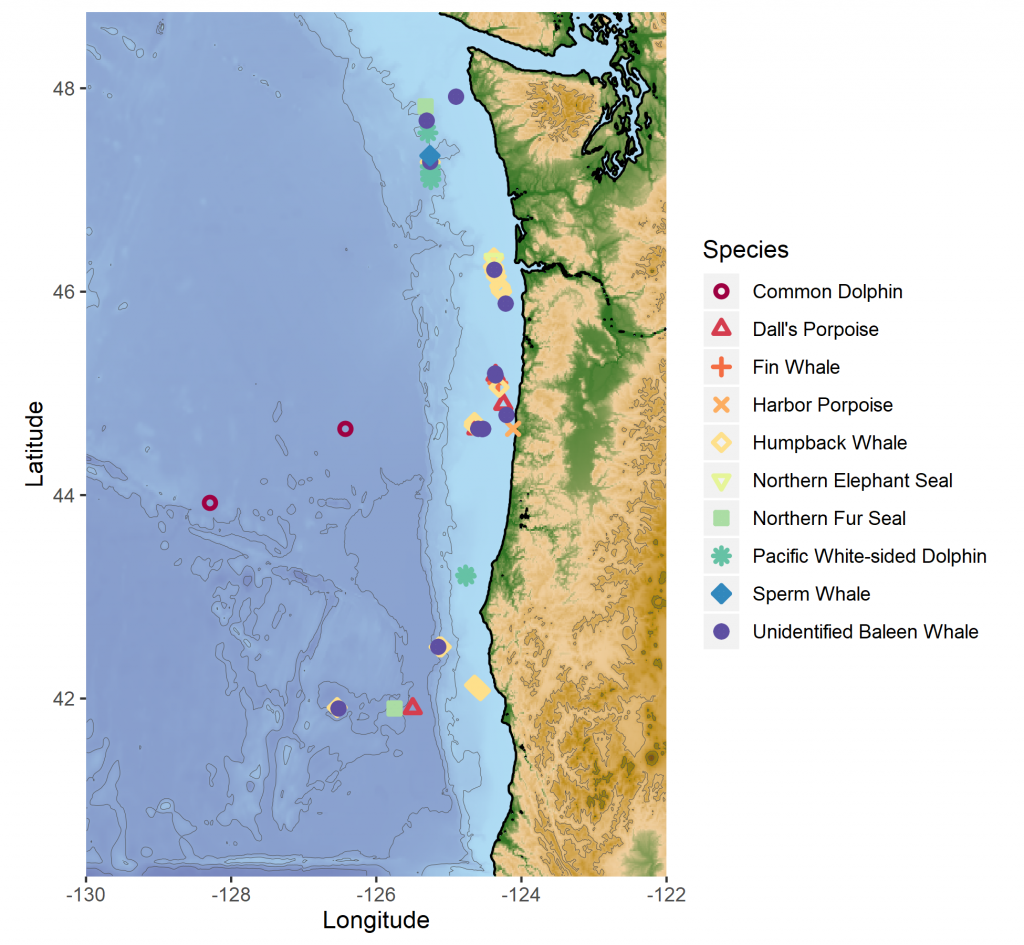
The fall 2019 NCC cruise was a resounding success. We were able to survey a large swath of the ecosystem between Crescent City, CA and La Push, WA, from inshore to 200 miles offshore. During that time, I observed nine different species of marine mammals (Table 1). As often as I use some version of the phrase “the marine environment is patchy and dynamic”, it never fails to sink in a little bit more every time I go to sea. On the map in Fig. 3, note how clustered the marine mammal sightings are. After nearly a full day of observing nothing but blue water, I would find myself scrambling to keep up with recording all the whales and dolphins we were suddenly in the midst of. What drives these clusters of sightings? What is it about the oceanography and prey community that makes any particular area a hotspot for marine mammals? We hope to get at these questions by utilizing the oceanographic data collected throughout the surveys to better understand environmental drivers of these distribution patterns.
Table 1. Summary of marine mammal
sightings from the September 2019 NCC Cruise.
Species
# sightings
Total # individuals
Northern Elephant Seal
1
1
Northern Fur Seal
2
2
Common Dolphin
2
8
Pacific White-sided Dolphin
8
143
Dall’s Porpoise
4
19
Harbor Porpoise
1
3
Sperm Whale
1
1
Fin Whale
1
1
Humpback Whale
22
36
Unidentified Baleen Whale
14
16
Figure 3. Map of marine mammal sighting locations from the September NCC cruise.
It was an auspicious time to survey the Northern California Current. Perhaps you have read recent news reports warning about the formation of another impending marine heatwave, much like the “warm blob” that plagued the North Pacific in 2015. We experienced it first-hand during the NCC cruise, with very warm surface waters off Newport extending out to 200 miles offshore (Fig. 4). A lot of energy input from strong winds would be required to mix that thick, warm layer and allow cool, nutrient-rich water to upwell along the coast. But it is already late September, and as the season shifts from summer to fall we are at the end of our typical upwelling season, and the north winds that would typically drive that mixing are less likely. Time will tell what is in store for the NCC ecosystem as we face the onset of another marine heatwave.
Figure 4. Temperature contours over the upper 150 m from 1-200 miles off Newport, Oregon from Fall 2014-2019. During Fall 2014, the Warm Blob inundated the Oregon shelf. Surface temperatures during that survey were 17°- 18°C along the entire transect. During 2015 and 2016 the warm water (16°C) layer had deepened and occupied the upper 50 m. During 2018, the temperature was 16°C in the upper 20 m and cooler on the shelf, indicative of residual upwelling. During this survey in 2019, we again saw very warm (18°C) temperatures in the upper water column over the entire transect. Image and caption credit: Jennifer Fisher.
It was a joy to spend 10 days at sea with this team of scientists. Insight, collaboration, and innovation are born from interdisciplinary efforts like the NCC cruises. Beyond science, what a privilege it is to be on the ocean with a group of people you can work with and laugh with, from the dock to 200 miles offshore, south to north and back again.
Dawn Barlow on the flying bridge of NOAA Ship Bell M. Shimada, heading out to sea with the Newport bridge in the background. Photo: Anna Bolm.
By Dominique Kone, Masters Student in Marine Resource Management
By now, I’m sure you’re aware of recent interests to reintroduce sea otters to Oregon. To inform this effort, my research focuses on predicting suitable sea otter habitat and investigating the potential ecological effects if sea otters are reintroduced in the future. This information will help managers gain a better understanding of the potential for sea otters to reestablish in Oregon, as well as how Oregon’s ecosystems may change via top-down processes. These analyses will address some sources of uncertainties of this effort, but there are still many more questions researchers could address to further guide this process. Here, I note some lingering questions I’ve come across in the course of conducting my research. This is not a complete list of all questions that could or should be investigated, but they represent some of the most interesting questions I have and others have in Oregon.
Credit: Todd Mcleish
The questions, and our associated knowledge on each of these topics:
Is there enough available prey to support a robust sea otter population in Oregon?
Sea otters require approximately 30% of their own body weight in food every day (Costa 1978, Reidman & Estes 1990). With a large appetite, they not only need to spend most of their time foraging, but require a steady supply of prey to survive. For predators, we assume the presence of suitable habitat is a reliable proxy for prey availability (Redfern et al. 2006). Whereby, quality habitat should supply enough prey to sustain predators at higher trophic levels.
In making these habitat predictions for sea otters, we must also recognize the potential limitations of this “habitat equals prey” paradigm, in that there may be parcels of habitat where prey is unavailable or inaccessible. In Oregon, there could be unknown processes unique to our nearshore ecosystems that would support less prey for sea otters. This possibility highlights the importance of not only understanding how much suitable habitat is available for foraging sea otters, but also how much prey is available in these habitats to sustain a viable otter population in the future. Supplementing these habitat predictions with fishery-independent prey surveys is one way to address this question.
Credit: Suzi Eszterhas via Smithsonian Magazine
How will Oregon’s oceanographic seasonality alter or impact habitat suitability?
Sea otters along the California coast exist in an environment with persistent Giant kelp beds, moderate to low wave intensity, and year-round upwelling regimes. These environmental variables and habitat factors create productive ecosystems that provide quality sea otter habitat and a steady supply of prey; thus, supporting high densities of sea otters. This environment contrasts with the Oregon coast, which is characterized by seasonal changes in bull kelp and wave intensity. Summer months have dense kelp beds, calm surf, and strong upwellings. While winter months have little to no kelp, weak upwellings, and intense wave climates. These seasonal variations raise the question as to how these temporal fluctuations in available habitat could impact the number of sea otters able to survive in Oregon.
In Washington – an environment like Oregon – sea otters exhibit seasonal distribution patterns in response to intensifying wave climates. During calm summer months, sea otters primarily forage along the outer coast, but move into more protected areas, such as the Strait of Juan de Fuca, during winter months (Laidre et al. 2009). If sea otters were reintroduced to Oregon, we may very well observe similar seasonal movement patterns (e.g. dispersal into estuaries), but the degree to which this seasonal redistribution and reduction in foraging habitat could impact sea otter reestablishment and recovery is currently unknown.
Credit: Oregon Coast Aquarium
In the event of a reintroduction, do northern or southern sea otters have a greater capacity to adapt to Oregon environments?
In the early 1970’s, Oregon’s first sea otter translocation effort failed (Jameson et al. 1982). Since then, hypotheses on the potential ecological differences between northern and southern sea otters have been proposed as potential factors of the failed effort, potentially due to different abilities to exploit specific prey species. Studies have demonstrated that northern and southern sea otters have slight morphological differences – northern otters having larger skulls and teeth than southern otters (Wilson et al. 1991). This finding has created the hypothesis that the northern otter’s larger skull and teeth allow it to consume prey with denser exoskeletons, and thereby can exploit a greater diversity of prey species. However, there appears to be a lack of evidence to suggest larger skulls and teeth translate to greater bite force. Based on morphology alone, either sub-species could be just as successful in exploiting different prey species.
A different direction to address questions around adaptability is to look at similarities in habitat and oceanographic characteristics. Sea otters exist along a gradient of habitat types (e.g. kelp forests, estuaries, soft-sediment environments) and oceanographic conditions (e.g. warm-temperature to cooler sub-Arctic waters) (Laidre et al. 2009, Lafferty et al. 2014). Yet, we currently don’t know how well or quickly otters can adapt when they expand into new habitats that differ from ones they are familiar with. Sea otters must be efficient foragers and need to acquire skills that allow them to effectively hunt specific prey species (Estes et al. 2003). Hypothetically, if we take sea otters from rocky environments where they’ve developed foraging skills to hunt sea urchins and abalones, and place them in a soft-sediment environment, how quickly would they develop new foraging skills to exploit soft-sediment prey species? Would they adapt quickly enough to meet their daily prey requirements?
Credit: Eric Risberg/Associated Press via The Columbian
In Oregon, specifically, how might climate change impact sea otters, and how might sea otters mediate climate impacts?
Climate change has been shown to directly impact many species via changes in temperature (Chen et al. 2011). Some species have specific thermal tolerances, in which they can only survive within a specified temperature range (i.e. maximum and minimum). Once the temperature moves out of that range, the species can either move with those shifting water masses, behaviorally adapt or perish (Sunday et al. 2012). It’s unclear if and how changing temperatures will impact sea otters, directly. However, sea otters could still be indirectly affected via impacts to their prey. If prey species in sea otter habitat decline due to changing temperatures, this would reduce available food for otters. Ocean acidification (OA) is another climate-induced process that could indirectly impact sea otters. By creating chemical conditions that make it difficult for species to form shells, OA could decrease the availability of some prey species, as well (Gaylord et al. 2011).
Interestingly, these pathways between sea otters and climate change become more complex when we consider the potentially mediating effects from sea otters. Aquatic plants – such as kelp and seagrass – can reduce the impacts of climate change by absorbing and taking carbon out of the water column (Krause-Jensen & Duarte 2016). This carbon sequestration can then decrease acidic conditions from OA and mediate the negative impacts to shell-forming species. When sea otters catalyze a tropic cascade, in which herbivores are reduced and aquatic plants are restored, they could increase rates of carbon sequestration. While sea otters could be an effective tool against climate impacts, it’s not clear how this predator and catalyst will balance each other out. We first need to investigate the potential magnitude – both temporal and spatial – of these two processes to make any predictions about how sea otters and climate change might interact here in Oregon.
Credit: National Wildlife Federation
In Summary
There are several questions I’ve noted here that warrant further investigation and could be a focus for future research as this potential sea otter reintroduction effort progresses. These are by no means every question that should be addressed, but they do represent topics or themes I have come across several times in my own research or in conversations with other researchers and managers. I think it’s also important to recognize that these questions predominantly relate to the natural sciences and reflect my interest as an ecologist. The number of relevant questions that would inform this effort could grow infinitely large if we expand our disciplines to the social sciences, economics, genetics, so on and so forth. Lastly, these questions highlight the important point that there is still a lot we currently don’t know about (1) the ecology and natural behavior of sea otters, and (2) what a future with sea otters in Oregon might look like. As with any new idea, there will always be more questions than concrete answers, but we – here in the GEMM Lab – are working hard to address the most crucial ones first and provide reliable answers and information wherever we can.
References:
Chen, I., Hill, J. K., Ohlemuller, R., Roy, D. B., and C. D. Thomas. 2011. Rapid range shifts of species associated with high levels of climate warming. Science. 333: 1024-1026.
Costa, D. P. 1978. The ecological energetics, water, and electrolyte balance of the California sea otter (Enhydra lutris). Ph.D. dissertation, University of California, Santa Cruz.
Estes, J. A., Riedman, M. L., Staedler, M. M., Tinker, M. T., and B. E. Lyon. 2003. Individual variation in prey selection by sea otters: patterns, causes and implications. Journal of Animal Ecology. 72: 144-155.
Gaylord et al. 2011. Functional impacts of ocean acidification in an ecologically critical foundation species. Journal of Experimental Biology. 214: 2586-2594.
Jameson, R. J., Kenyon, K. W., Johnson, A. M., and H. M. Wight. 1982. History and status of translocated sea otter populations in North America. Wildlife Society Bulletin. 10(2): 100-107.
Krause-Jensen, D., and C. M. Duarte. 2016. Substantial role of macroalgae in marine carbon sequestration. Nature Geoscience. 9: 737-742.
Lafferty, K. D., and M. T. Tinker. 2014. Sea otters are recolonizing southern California in fits and starts. Ecosphere.5(5).
Laidre, K. L., Jameson, R. J., Gurarie, E., Jeffries, S. J., and H. Allen. 2009. Spatial habitat use patterns of sea otters in coastal Washington. Journal of Marine Mammalogy. 90(4): 906-917.
Redfern et al. 2006. Techniques for cetacean-habitat modeling. Marine Ecology Progress Series. 310: 271-295.
Reidman, M. L. and J. A. Estes. 1990. The sea otter (Enhydra lutris): behavior, ecology, and natural history. United States Department of the Interior, Fish and Wildlife Service, Biological Report. 90: 1-126.
Sunday, J. M., Bates, A. E., and N. K. Dulvy. 2012. Thermal tolerance and the global redistribution of animals. Nature: Climate Change. 2: 686-690.
Wilson, D. E., Bogan, M. A., Brownell, R. L., Burdin, A. M., and M. K. Maminov. 1991. Geographic variation in sea otters, Ehydra lutris. Journal of Mammalogy. 72(1): 22-36.
By Christina Garvey, University of Maryland, GEMM Lab REU Intern
It is July 8th and it is my 4th week here in Hatfield as an REU intern for Dr. Leigh Torres. My name is Christina Garvey and this summer I am studying the spatial ecology of blue whales in the South Taranaki Bight, New Zealand. Coming from the east coast, Oregon has given me an experience of a lifetime – the rugged shorelines continue to take my breath away and watching sea lions in Yaquina Bay never gets old. However, working on my first research project has by far been the greatest opportunity and I have learned so much in so little time. When Dr. Torres asked me to contribute to this blog I was unsure of how I would write about my work thus far but I am excited to have the opportunity to share the knowledge I have gained with whoever reads this blog post.
The research project that I will be conducting this summer will use remotely sensed environmental data (information collected from satellites) to predict blue whale distribution in the South Taranaki Bight (STB), New Zealand. Those that have read previous blogs about this research may remember that the STB study area is created by a large indentation or “bight” on the southern end of the Northern Island. Based on multiple lines of evidence, Dr. Leigh Torres hypothesized the presence of an unrecognized blue whale foraging ground in the STB (Torres 2013). Dr. Torres and her team have since proved that blue whales frequent this region year-round; however, the STB is also very industrial making this space-use overlap a conservation concern (Barlow et al. 2018). The increasing presence of marine industrial activity in the STB is expected to put more pressure on blue whales in this region, whom are already vulnerable from the effects of past commercial whaling (Barlow et al. 2018) If you want to read more about blue whales in the STB check out previous blog posts that talk all about it!
Figure 1. A blue whale surfaces in front of a floating production storage and offloading vessel servicing the oil rigs in the South Taranaki Bight. Photo by D. Barlow.
Figure 2. South Taranaki Bight, New Zealand, our study site outlined by the red box. Kahurangi Point (black star) is the site of wind-driven upwelling system.
The possibility of the STB as an important foraging ground for a resident population of blue whales poses management concerns as New Zealand will have to balance industrial growth with the protection and conservation of a critically endangered species. As a result of strong public support, there are political plans to implement a marine protected area (MPA) in the STB for the blue whales. The purpose of our research is to provide scientific knowledge and recommendations that will assist the New Zealand government in the creation of an effective MPA.
In order to create an MPA that would help conserve the blue whale population in the STB, we need to gather a deeper understanding of the relationship between blue whales and this marine environment. One way to gain knowledge of the oceanographic and ecological processes of the ocean is through remote sensing by satellites, which provides accessible and easy to use environmental data. In our study we propose remote sensing as a tool that can be used by managers for the design of MPAs (through spatial and temporal boundaries). Satellite imagery can provide information on sea surface temperature (SST), SST anomaly, as well as net primary productivity (NPP) – which are all measurements that can help describe oceanographic upwelling, a phenomena that is believed to be correlated to the presence of blue whales in the STB region.
Figure 3. The stars of the show: blue whales. A photograph captured from the small boat of one animal fluking up to dive down as another whale surfaces close by. (Photo credit: L. Torres)
Past studies in the STB showed evidence of a large upwelling event that occurs off the coast of Kahurangi Point (Fig. 2), on the northwest tip of the South Island (Shirtcliffe et al. 1990). In order to study the relationship of this upwelling to the distribution of blue whales, I plan to extract remotely sensed data (SST, SST anomaly, & NPP) off the coast of Kahurangi and compare it to data gathered from a centrally located site within the STB, which is close to oil rigs and so is of management interest. I will first study how decreases in sea surface temperature at the site of upwelling (Kahurangi) are related to changes in sea surface temperature at this central site in the STB, while accounting for any time differences between each occurrence. I expect that this relationship will be influenced by the wind patterns, and that there will be changes based on the season. I also predict that drops in temperature will be strongly related to increases in primary productivity, since upwelling brings nutrients important for photosynthesis up to the surface. These dips in SST are also expected to be correlated to blue whale occurrence within the bight, since blue whale prey (krill) eat the phytoplankton produced by the productivity.
Figure 4. A blue whale lunges on an aggregation of krill. UAS piloted by Todd Chandler.
To test the relationships I determine between remotely sensed data at different locations in the STB, I plan to use blue whale observations from marine mammal observers during a seismic survey conducted in 2013, as well as sightings recorded from the 2014, 2016, and 2017 field studies led by Dr. Leigh Torres. By studying the statistical relationships between all of these variables I hope to prove that remote sensing can be used as a tool to study and understand blue whale distribution.
I am very excited about this research, especially because the end goal of creating an MPA really gives me purpose. I feel very lucky to be part of a project that could make a positive impact on the world, if only in just a little corner of New Zealand. In the mean time I’ll be here in Hatfield doing the best I can to help make that happen.
References:
Barlow DR, Torres LG, Hodge KB, Steel D, Baker CS, Chandler TE, Bott N, Constantine R, Double MC, Gill P, Glasgow D, Hamner RM, Lilley C, Ogle M, Olson PA, Peters C, Stockin KA, Tessaglia-hymes CT, Klinck H (2018) Documentation of a New Zealand blue whale population based on multiple lines of evidence. Endanger Species Res 36:27–40.
Shirtcliffe TGL, Moore MI, Cole AG, Viner AB, Baldwin R, Chapman B (1990) Dynamics of the Cape Farewell upwelling plume, New Zealand. New Zeal J Mar Freshw Res 24:555–568.
Torres LG (2013) Evidence for an unrecognised blue whale foraging ground in New Zealand. New Zeal J Mar Freshw Res 47:235–248.
By: Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
For the past two years, I’ve had the opportunity to be the marine mammal observer aboard the NOAA ship Bell M. Shimada for 10 days in May. Both trips covered transects in the Northern California Current Ecosystem during the same time of year, but things looked very different from my chair on the fly bridge. This trip, in particular, highlighted the importance of standardization, seeing as it was the second replicate of the same area. Other scientists and crew members repeatedly asked me the same questions that made me realize just how important it is to have standards in scientific practices and communicating them.
Northern right whale dolphin porpoising out of the water beside the ship while in transit. May 2019. Image source: Alexa Kownacki
The questions:
What do you actually do here and why are you doing it?
Is this year the same as last year in terms of weather, sightings, and transect locations?
Did you expect to see greater or fewer sightings (number and diversity)?
What is this Beaufort Sea State scale that you keep referring to?
All of these are important scientific questions that influence our hypothesis-testing research, survey methods, expected results, and potential conclusions. Although the entire science party aboard the ship conducted marine science, we all had our own specialties and sometimes only knew the basics, if that, about what the other person was doing. It became a perfect opportunity to share our science and standards across similar, but different fields.
Now, to answer those questions:
a) What do you actually do here and b) why are you doing it?
a) As the only marine mammal observer, I stand watch during favorable weather conditions while the ship is in transit, scanning from 0 to 90 degrees off the starboard side (from the front of the ship to a right angle towards the right side when facing forwards). Meanwhile, an application on an iPad called SeaScribe, records the ship’s exact location every 15 seconds, even when no animal is sighted. This process allows for the collection of absence data, that is, data when no animals are present. The SeaScribe program records the survey lines, along with manual inputs that I add, including weather and observer information. When I spot a marine mammal, I immediately mark an exact location on a hand held GPS, use my binoculars to identify the species, and add information to the sighting on the SeaScribe program, such as species, distance to the sighted animal(s), the degree (angle) to the sighting, number of animals in a group, behavior, and direction if traveling.
b) Marine mammal observing serves many different purposes. In this case, observing collects information about what species are where at what time. By piggy-backing on these large-scale, offshore oceanographic NOAA surveys, we have the unique opportunity to survey along standardized transect lines during different times of the year. From replicate survey data, we can start to form an idea of which species use which areas and what oceanographic conditions may impact species distributions. Currently there is not much consistent marine mammal data collected over these offshore areas between Northern California and Washington State, so our work is aiming to fill this knowledge gap.
Alexa observing on the R/V Shimada in May 2019, all bundled up. Image Source: Alexa Kownacki
What is this Beaufort Sea State scale that you keep referring to?
Great question! It took me a while to realize that this standard measuring tool to estimate wind speeds and sea conditions, is not commonly recognized even among other sea-goers. The Beaufort Sea State, or BSS, uses an empirical scale that ranges from 0-12 with 0 being no wind and calm seas, to 12 being hurricane-force winds with 45+ ft seas. It is frequently referenced by scientists in oceanography, marine science, and climate science as a universally-understood metric. The BSS was created in 1805 by Francis Beaufort, a hydrographer in the Royal Navy, to standardize weather conditions across the fleet of vessels. By the mid-1850s, the BSS was standardized to non-naval use for sailing vessels, and in 1916, expanded to include information specific to the seas and not the sails1. We in the marine mammal observation field constantly collect BSS information while on survey to measure the quality of survey conditions that may impact our observations. BSS data allows us to measure the extent of our survey range, both in the distance that we are likely to sight animals and also the likelihood of sighting anything. Therefore, the BSS scale gives us an important indication of how much absence data we have collected, in addition to presence data.
A description of the Beaufort Sea State Scale. Image source: National Weather Service.
Is this year the same as last year in terms of weather, sightings, and transect locations?
The short answer is no. Observed differences in marine mammal sightings in terms of both species diversity and number of animals between years can be normal. There are many potential explanatory variables, from differences in currents, upwelling strength, El Nino index levels, water temperatures, or, what was obvious in this case: sighting conditions. The weather in May 2019 varied greatly from that in May 2018. Last year, I observed for nearly every day because the Beaufort Sea State (BSS) was frequently less than a four. However, this year, more often than not, the BSS greater than or equal to five. A BSS of 5 equates to approximately 17-21 knots of breeze with 6-foot waves and the water appears to have many “white horses” or pronounced white caps with sea spray. Additionally, mechanical issue with winches delayed and altered our transect locations. Therefore, although multiple transects from May 2018 were also surveyed during May 2019, there were a few lines that do not have data for both cruises.
May 2018 with a BSS 1
May 2019 with a BSS 6
Did you expect to see greater or fewer sightings (number and diversity)?
Knowing that I had less favorable sighting conditions and less amount of effort observing this year, it is not surprising that I observed fewer marine mammals in total count and in species diversity. Even less surprising is that on the day with the best weather, where the BSS was less than a five, I recorded the most sightings with the highest species count. May 2018 felt a bit like a tropical vacation because we had surprisingly sunny days with mild winds, and during May 2019 we had some rough seas with gale force winds. Additionally, as an observer, I need to remove as much bias as possible. So, yes, I had hoped to see beaked whales or orca like I did in May 2018, but I was still pleasantly surprised when I spotted fin whales feeding in May 2019.
Standardization is a common theme. Observing between years on standard transects, at set speeds, in different conditions using standardized tools is critical to collecting high quality data that is comparable across different periods. Scientists constantly think about quality control. We look for trends and patterns, similarities and differences, but none of those could be understood without having standard metrics.
The entire science party aboard the R/V Shimada in May 2019, including a marine mammal scientist, phytoplankton scientists, zooplankton scientists, and fisheries scientists, and oceanographers. Image Source: Alexa Kownacki
Literature Cited:
1Oliver, John E. (2005). Encyclopedia of world climatology. Springer.
I have the privilege of studying the largest animals on the planet: blue whales (Balaenoptera musculus). However, in order to understand the ecology, distribution, and habitat use patterns of these ocean giants, I have dedicated the past several months to studying something much smaller: krill (Nyctiphanes australis). New Zealand’s South Taranaki Bight region (“STB”, Figure 1) is an important foraging ground for a unique population of blue whales [1,2]. A wind-driven upwelling system off of Kahurangi Point (the “X” in Figure 1) generates productivity in the region [3], leading to an abundance of krill [4], the desired blue whale prey [5].
Figure 1. A map of New Zealand, with the South Taranaki Bight (STB) region delineated by the black box. Cape Farewell is denoted by a star, and Kahurangi point is denoted by an X.
Figure 4. A blue whale lunges on an aggregation of krill. UAS piloted by Todd Chandler.
Krill (Nyctiphanes australis), blue whale prey in New Zealand’s South Taranaki Bight region. Image: Te Papa.
Our blue whale research team collected a multitude of datastreams in three different years, including hydroacoustic data to map krill distribution throughout our study region. The summers of 2014 and 2017 were characterized by what could be considered “typical” conditions: A plume of cold, upwelled water curving its way around Cape Farewell (marked with the star in Figure 1) and entering the South Taranaki Bight, spurring a cascade of productivity in the region. The 2016 season, however, was different. The surface water temperatures were hot, and the whales were not where we expected to find them.
Figure 2. Sea surface temperature maps of the South Taranaki Bight region in each of our three study years. The white circles indicate where most blue whale sightings were made in each year. Note the very warm temperatures in 2016, and more westerly location of blue whale sightings.
What happened to the blue whales’ food source under these different conditions in 2016? Before I share some preliminary findings from my recent analyses, it is important to note that there are many possible ways to measure krill availability. For example, the number of krill aggregations, as well as how deep, thick, and dense those aggregations are in an area will all factor into how “desirable” krill patches are to a blue whale. While there may not be “more” or “less” krill from one year to the next, it may be more or less accessible to a blue whale due to energetic costs of capturing it. Here is a taste of what I’ve found so far:
In 2016, when surface waters were warm, the krill aggregations were significantly deeper than in the “typical” years (ANOVA, F=7.94, p <0.001):
Figute 3. Boxplots comparing the median krill aggregation depth in each of our three survey years.
The number of aggregations was not significantly different between years, but as you can see in the plot below (Figure 4) the krill were distributed differently in space:
Figure 4. Map of the South Taranaki Bight region with the number of aggregations per 4 km^2, standardized by vessel survey effort. The darker colors represent areas with a higher density of krill aggregations.
While the bulk of the krill aggregations were located north of Cape Farewell under typical conditions (2014 and 2017), in the warm year (2016) the krill were not in this area. Rather, the area with the most aggregations was offshore, in the western portion of our study region. Now, take a look at the same figure, overlaid with our blue whale sighting locations:
Figure 5. Map of standardized number of krill aggregations, overlaid with blue whale sighting locations in red stars.
Where did we find the whales? In each year, most whale encounters were in the locations where the most krill aggregations were found! Not only that, but in 2016 the whales responded to the difference in krill distribution by shifting their distribution patterns so that they were virtually absent north of Cape Farewell, where most sightings were made in the typical years.
The above figures demonstrate the importance of studying an ecosystem. We could puzzle and speculate over why the blue whales were further west in the warm year, but the story that is emerging in the krill data may be a key link in our understanding of how the ecosystem responds to warm conditions. While the focus of my dissertation research is blue whales, they do not live in isolation. It is through understanding the ecosystem-scale story that we can better understand blue whale ecology in the STB. As I continue modeling the relationships between oceanography, krill, and blue whales in warm and typical years, we are beginning to scratch the surface of how blue whales may be responding to their environment.
Torres LG. 2013 Evidence for an unrecognised blue whale foraging ground in New Zealand. New Zeal. J. Mar. Freshw. Res.47, 235–248. (doi:10.1080/00288330.2013.773919)
Barlow DR et al. 2018 Documentation of a New Zealand blue whale population based on multiple lines of evidence. Endanger. Species Res.36, 27–40. (doi:https://doi.org/10.3354/esr00891)
Shirtcliffe TGL, Moore MI, Cole AG, Viner AB, Baldwin R, Chapman B. 1990 Dynamics of the Cape Farewell upwelling plume, New Zealand. New Zeal. J. Mar. Freshw. Res.24, 555–568. (doi:10.1080/00288330.1990.9516446)
Bradford-Grieve JM, Murdoch RC, Chapman BE. 1993 Composition of macrozooplankton assemblages associated with the formation and decay of pulses within an upwelling plume in greater cook strait, New Zealand. New Zeal. J. Mar. Freshw. Res.27, 1–22. (doi:10.1080/00288330.1993.9516541)
Gill P. 2002 A blue whale (Balaenoptera musculus) feeding ground in a southern Australian coastal upwelling zone. J. Cetacean Res. Manag.4, 179–184.
By Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
As much as I try to keep politics out of my science vocabulary, there are some ties between the two that cannot be severed. Often, science in the United States is very linked to the government because funding can be dependent on federal, state, and/or local government decisions. Therefore, it is part of our responsibility as scientists to be, at least, informed on governmental proceedings.
The United States has one agency that is particularly important to those of us conducting marine science: the National Oceanic and Atmospheric Administration (NOAA). NOAA’s mission is science, service, and stewardship with three major components:
To understand and predict changes in climate, weather, oceans and coasts
To share that knowledge and information with others
To conserve and manage coastal and marine ecosystems and resources
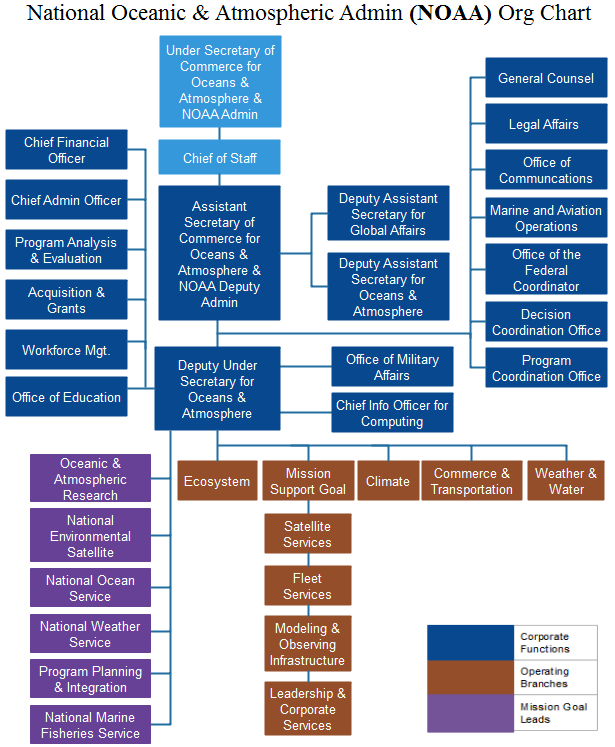
Organizational Chart of NOAA. (Image source: OrgCharting)
Last year, the U.S. Senate confirmed Retired Rear Admiral Timothy Gallaudet, Ph.D., as the Assistant Secretary of Commerce for Oceans and Atmosphere for the Department of Commerce in NOAA. This position is an appointment by the current President of the United States, and is tasked with overseeing the daily functions and the strategic and operational future of NOAA. NOAA oversees the National Marine Fisheries Service (NMFS), which is an agency responsible for the stewardship and management of the nation’s living marine resources. NMFS is a major player when it comes to marine science, particularly through the determination of priorities for research and management of marine species and habitats within the United States’ exclusive economic zone (EEZ).
In dark blue, the United States’ Exclusive Economic zones, surrounding land masses in green. (Figure by K. Laws)
Recently, I had the opportunity to hear Dr. Gallaudet speak to scientists who work for, or in conjunction with, a NMFS office. After the 16% budget cut from the fiscal year 2017 to 2018, many marine scientists are concerned about how budget changes will impact research. Therefore, I knew Dr. Gallaudet’s visit would provide insight about the future of marine science in the United States.
Dr. Gallaudet holds master’s and doctoral degrees in oceanography from Scripps Institution of Oceanography, as well as a bachelor’s degree from the United States Naval Academy. He spent 32 years in the Navy before stepping into his current role as Assistant Secretary. Throughout the meeting, Dr. Gallaudet emphasized his leadership motto: All in, All Good, and All for One.
Dr. Gallaudet also spoke about where he sees NOAA moving towards: the private sector.
A prominent conservation geneticist asked Dr. Gallaudet how NOAA can better foster advanced degree-seeking students. The geneticist commented that a decade ago there were 10-12 PhD students in this one science center alone. Today, there is “maybe one”. Dr. Gallaudet responded that the science centers should start reaching out to private industry. In response to other questions, he continued to redirect scientists toward United States-based corporations that could join forces with government agencies. He believes that if NMFS scientists share data and projects with local biotechnology, medical, and environmental companies, the country can foster positive relationships with industry. Dr. Gallaudet commented that the President wants to create these win-win situations: where the US government pairs with for-profit companies. It is up to us, as the scientists, how we make those connections.
As scientists, we frequently avoid heated political banter in the hopes of maintaining an objective and impartial approach to our research. However, these lines can be blurred. Much of our science depends on political decisions that mold our future, including how funding is allocated and what goals are prioritized. In 2010, Science Magazine published an online article, “Feeding your Research into the Policy Debate” where Elisabeth Pain highlighted the interdisciplinary nature of science and policy. In Pain’s interview with Troy Benn, a PhD student in Urban Ecology at the time, Benn comments that he learned just how much scientists play a role in policy and how research contributes to policy deliberations. Sometimes our research becomes of interest to politicians and sometimes it is the other way around.
From my experiences collaborating with government entities, private corporations, and nonprofit organizations, I realize that science-related policy is imperative. California established a non-profit, the California Ocean Science Trust (OST), for the specific objective supporting management decisions with the best science and bridging science and policy. A critical analysis of the OST by Pietri et al., “Using Science to Inform Controversial Issues: A Case Study from the California Ocean Science Trust”, matches legislation with science. For example, the Senate Bill (SB) 1319, better known as the California Ocean Protection Act (COPA), calls for “decisions informed by good science” and to “advance scientific understanding”. Science is explicitly written into legislation and I think that is a call to action. If an entire state can mobilize resources to create a team of interdisciplinary experts, I can inform myself on the politics that have potential to shape my future and the future of my science.
An image of the NOAA ship Bell M. Shimada transiting between stations. Multiple members of the GEMM Lab conducted surveys from this NOAA vessel in 2018. (Image source: Alexa Kownacki)
By Dawn Barlow, PhD student, Department of Fisheries & Wildlife, Geospatial Ecology of Marine Megafauna Lab
When people hear that I study blue whales, they often ask me questions about what it’s like to be close to the largest animal on the planet, where we do fieldwork, and what data we are interested in collecting. While I love time at sea, my view on a daily basis is rarely like this:
Our small research vessel at sunset in New Zealand’s South Taranaki Bight at the end of a day of blue whale survey. Photo by D. Barlow.
More often than not, it looks something like this:
In my application letter to Dr. Leigh Torres, I wrote something along the lines of “while I relish remote fieldwork, I also find great satisfaction in the analysis process.” This statement is increasingly true for me as I grow more proficient in statistical modeling and computer programming. When excitedly telling my family about how I am trying to model relationships between oceanography, krill, whales, and satellite imagery, I was asked what I meant by “model”. Put simply, a model is a formula or equation that we can use to describe a pattern. I have been told, “all models are wrong, but some models work.” What does this mean? While we may never know exactly every pattern of whale feeding behavior, we can use the data we have to describe some of the important relationships. If our model performance is very good, then we have likely described most of what drives the patterns we see. If model performance is poor, then there is more to the pattern that we have not yet captured in either our data collection or in our analytical methods. Another common saying about models is, “A model is only ever as good as the data you put into it.” While we worked hard during field seasons to collect a myriad of data about what could be influencing blue whale distribution patterns, we inevitably could not capture everything, nor do we know everything that should be measured.
So, how do you go about finding the ‘best’ model? This question is what I’ve been grappling with over the last several weeks. My goal is to describe the patterns in the krill that drive patterns in whale distribution, the patterns in oceanography that drive patterns in the krill, and the patterns in the oceanography that drive patterns in whale distribution. The thing is, we have many metrics to describe oceanographic patterns (surface temperature, mixed layer depth, strength of the thermocline, integral of fluorescence, to name just a few), as well as several metrics to describe the krill (number of aggregations, aggregation density, depth, and thickness). When I multiplied out how many possible combinations of predictor variables and parameters we’re interested in modeling, I realized this meant running nearly 300 models in order to settle on the best ten. This is where programming comes in, I told myself, and caught my breath.
I’ve always loved languages. When I was much younger, I thought I might want to study linguistics. As a graduate student in wildlife science, the language I’ve spent the most time learning, and come to love, is the statistical programming language R. Just like any other language, R has syntax and structure. Like any other language, there are many ways in which to articulate something, to make a particular point or reach a particular end goal. Well-written code is sometimes described as “elegant”, much like a well-articulated piece of writing. While I certainly do not consider myself “fluent” in R, it is a language I love learning. I like to think that the R scripts I write are an attempt to eloquently uncover and describe ecological patterns.
Rather than running 300 models one by one, I wrote an R script to run many models at a time, and then sort the outputs by model performance. I may look at the five best models of 32 options in order to select one. But this is where Leigh reminds me to step back from the programming for a minute and put my ecologist hat back on. Insight on the part of the modeler is needed in order to discern between what are real ecological relationships and what are spurious correlations in the data. It may not be quite as simple as choosing the model with the highest explanatory power when my goal is to make ecological inferences.
So, where does this leave me? Hundreds of models later, I am still not entirely sure which ones are best, although I’ve narrowed it down considerably. My programming proficiency and confidence continue to grow, but that only goes so far in ecology. Knowledge of my study system is equally important. So my workflow lately goes something like this: write code, try to interpret model outputs, consider what I know about the oceanography of my study region, re-write code, re-interpret the revised results, and so on. Hopefully this iterative process is bringing us gradually closer to an understanding of the ecology of blue whales on a foraging ground… stay tuned.
A blue whale lunges on an aggregation of krill in New Zealand’s South Taranaki Bight. Drone piloted by Todd Chandler.
By Dawn Barlow, PhD student, Geospatial Ecology of Marine Megafauna Lab, Department of Fisheries and Wildlife, Oregon State University
Sun on my face and wind in my hair, scanning the expanse of blue. Forty minutes on, twenty minutes off, from sunrise until sunset, day after day. Hours of seemingly empty blue, punctuated by graceful black-footed albatrosses wheeling and gliding over the swells, by the splashing approach of a curious group of Pacific white-sided dolphins coming to play in the bow of the ship, by whale spouts on the horizon and the occasional breaching humpback. A flurry of data entry—geographic coordinates, bearing and distance from the ship, number of animals, species identification, behavior—and then back to blue.
Scanning for marine mammals from the flying bridge of NOAA ship Bell M. Shimada. Photo: Jess O’Loughlin.
I’ve just returned from the Northern California Current (NCC) ecosystem cruise aboard NOAA ship Bell M. Shimada. My role on board was the marine mammal observer, logging marine mammal sightings during the transits between sampling stations. We surveyed and sampled between Cape Mears, Oregon and Trinidad, California, from right along the coast out to 200 nautical miles offshore. Resources in the marine environment are patchy, and our coastline is highly productive. This diversity in environmental conditions creates niche habitats for many species, which is one reason why surveying and sampling across a broad geographic range can be so informative. We left Newport surrounded by gray whales, feeding in green, chilly waters at temperatures around 12°C. Moving west, the marine mammal and seabird sightings were increasingly sparse, the water increasingly blue, and the surface temperature warmed to a balmy 17°C. We had reached offshore waters, an ocean region sometimes referred to as the “blue desert”. For an entire day I didn’t see a single marine mammal and only just a few seabirds, until a handful of common dolphins—more frequently seen in warm-temperate and tropical waters to the south—joined the ship at sunset. As we transited back inshore over the productive Heceta Bank, the water became cooler and greener. I stayed busy logging sightings of humpback and gray whales, harbor porpoise and Dall’s porpoise, pacific white-sided dolphins and sea lions. These far-ranging marine predators must find a way to make a living in the patchy and dynamic ocean environment, and therefore their distribution is also patchy—aggregated around areas of high productivity and prey availability, and occasionally seen transiting in between.
Here are a few cruise highlights:
Curious groups of common dolphins (Delphinus delphis) came to play in the bow wake of the ship and even checked out the plankton nets when they were deployed. Common dolphins are typically found further south, however we saw several groups of them in the warmer waters far offshore.
Common dolphins (Delphinus delphis). Photo: Dawn Barlow.
Common dolphins (Delphinus delphis). Photo: Dawn Barlow.
Common dolphins (Delphinus delphis). Photo: Dawn Barlow.
Common dolphins (Delphinus delphis). Photo: Dawn Barlow.
Ocean sunfish (Mola mola) will occasionally lay themselves flat at the surface so that seabirds will pick them clean of any parasites. I was delighted to observe this for the first time just off Newport! There were several more sunfish sightings throughout the cruise.
Gull picking parasites off an ocean sunfish (Mola mola). Photo: Dawn Barlow.
A masked booby (Sula dactylatra) hung around the ship for a bit, 16 nautical miles from shore, just south of the Oregon-California border. Considered a tropical species, a sighting this far north is extremely rare. While masked boobies are typically distributed in the Caribbean and tropical Pacific from Mexico to Australia, one found its way to the Columbia River in 2006 (first record in the state of Oregon) and another showed up here to Newport in 2015 – reportedly only the second to be recorded north of Mendocino County, California. Perhaps this sighting is the third?
Masked booby (Sula dactylatra). Photo: Dawn Barlow.
While most of my boat-based fieldwork experiences have been focused on marine mammal research, this was an interdisciplinary cruise aimed at studying multiple aspects of the northern California current ecosystem. There were researchers on board studying oceanography, phytoplankton and harmful algal blooms, zooplankton, and microplastics. When a group of enthusiastic scientists with different areas of expertise come together and spend long days at sea, there is a wonderful opportunity to learn from one another. The hydroacoustic backscatter on the scientific echosounder prompted a group discussion about vertical migration of plankton one evening. Another evening I learned about differences in energetic content between krill species, and together we mused about what that might mean for marine predators. This is how collaborations are born, and I am grateful for the scientific musings with so many insightful people.
Thank you to the Shimada crew and the NCC science team for a wonderful cruise!
By Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
Did you know that Excel has a maximum number of rows? I do. During Winter Term for my GIS project, I was using Excel to merge oceanographic data, from a publicly-available data source website, and Excel continuously quit. Naturally, I assumed I had caused some sort of computer error. [As an aside, I’ve concluded that most problems related to technology are human error-based.] Therefore, I tried reformatting the data, restarting my computer, the program, etc. Nothing. Then, thanks to the magic of Google, I discovered that Excel allows no more than 1,048,576 rows by 16,384 columns. ONLY 1.05 million rows?! The oceanography data was more than 3 million rows—and that’s with me eliminating data points. This is what happens when we’re dealing with big data.
According to Merriam-Webster dictionary, big data is an accumulation of data that is too large and complex for processing by traditional database management tools (www.merriam-webster.com). However, there are journal articles, like this one from Forbes, that discuss the ongoing debate of how to define “big data”. According to the article, there are 12 major definitions; so, I’ll let you decide what you qualify as “big data”. Either way, I think that when Excel reaches its maximum row capacity, I’m working with big data.
Collecting oceanography data aboard the R/V Shimada. Photo source: Alexa K.
Here’s the thing: the oceanography data that I referred to was just a snippet of my data. Technically, it’s not even MY data; it’s data I accessed from NOAA’s ERDDAP website that had been consistently observed for the time frame of my dolphin data points. You may recall my blog about maps and geospatial analysis that highlights some of the reasons these variables, such as temperature and salinity, are important. However, what I didn’t previously mention was that I spent weeks working on editing this NOAA data. My project on common bottlenose dolphins overlays environmental variables to better understand dolphin population health off of California. These variables should have similar spatiotemporal attributes as the dolphin data I’m working with, which has a time series beginning in the 1980s. Without taking out a calculator, I still know that equates to a lot of data. Great data: data that will let me answer interesting, pertinent questions. But, big data nonetheless.
This is a screenshot of what the oceanography data looked like when I downloaded it to Excel. This format repeats for nearly 3 million rows.
Excel Screen Shot. Image source: Alexa K.
I showed this Excel spreadsheet to my GIS professor, and his response was something akin to “holy smokes”, with a few more expletives and a look of horror. It was not the sheer number of rows that shocked him; it was the data format. Nowadays, nearly everyone works with big data. It’s par for the course. However, the way data are formatted is the major split between what I’ll call “easy” data and “hard” data. The oceanography data could have been “easy” data. It could have had many variables listed in columns. Instead, this data alternated between rows with variable headings and columns with variable headings, for millions of cells. And, as described earlier, this is only one example of big data and its challenges.
Data does not always come in a form with text and numbers; sometimes it appears as media such as photographs, videos, and audio files. Big data just got a whole lot bigger. While working as a scientist at NOAA’s Southwest Fisheries Science Center, one project brought in over 80 terabytes of raw data per year. The project centered on the eastern north pacific gray whale population, and, more specifically, its migration. Scientists have observed the gray whale migration annually since 1994 from Piedras Blancas Light Station for the Northbound migration, and 2 out of every 5 years from Granite Canyon Field Station (GCFS) for the Southbound migration. One of my roles was to ground-truth software that would help transition from humans as observers to computer as observers. One avenue we assessed was to compare how well a computer “counted” whales compared to people. For this question, three infrared cameras at the GCFS recorded during the same time span that human observers were counting the migratory whales. Next, scientists, such as myself, would transfer those video files, upwards of 80 TB, from the hard drives to Synology boxes and to a different facility–miles away. Synology boxes store arrays of hard drives and that can be accessed remotely. To review, three locations with 80 TB of the same raw data. Once the data is saved in triplet, then I could run a computer program, to detect whale. In summary, three months of recorded infrared video files requires upwards of 240 TB before processing. This is big data.
Scientists on an observation shift at Granite Canyon Field Station in Northern California. Photo source: Alexa K.
Alexa and another NOAA scientist watching for gray whales at Piedras Blancas Light Station. Photo source: Alexa K.
In the GEMM Laboratory, we have so many sources of data that I did not bother trying to count. I’m entering my second year of the Ph.D. program and I already have a hard drive of data that I’ve backed up three different locations. It’s no longer a matter of “if” you work with big data, it’s “how”. How will you format the data? How will you store the data? How will you maintain back-ups of the data? How will you share this data with collaborators/funders/the public?
The wonderful aspect to big data is in the name: big and data. The scientific community can answer more, in-depth, challenging questions because of access to data and more of it. Data is often the limiting factor in what researchers can do because increased sample size allows more questions to be asked and greater confidence in results. That, and funding of course. It’s the reason why when you see GEMM Lab members in the field, we’re not only using drones to capture aerial images of whales, we’re taking fecal, biopsy, and phytoplankton samples. We’re recording the location, temperature, water conditions, wind conditions, cloud cover, date/time, water depth, and so much more. Because all of this data will help us and help other scientists answer critical questions. Thus, to my fellow scientists, I feel your pain and I applaud you, because I too know that the challenges that come with big data are worth it. And, to the non-scientists out there, hopefully this gives you some insight as to why we scientists ask for external hard drives as gifts.
Leila launching the drone to collect aerial images of gray whales to measure body condition. Photo source: Alexa K.
Using the theodolite to collect tracking data on the Pacific Coast Feeding Group in Port Orford, OR. Photo source: Alexa K.
By Dawn Barlow, PhD student, Department of Fisheries & Wildlife, Geospatial Ecology of Marine Megafauna Lab
As a PhD student studying the ecology of blue whales in New Zealand, my time is occupied by questions such as: When and where are the blue whales? Can we predict where they will be based on environmental conditions? How does their distribution overlap with human activity such as oil and gas exploration?
Leigh and I have just returned from New Zealand, where I gave an oral presentation at the Society for Conservation Biology Oceania Congress entitled “Cloudy with a chance of whales: Forecasting blue whale presence to mitigate industrial impacts based on tiered, bottom-up models”. While the findings I presented are preliminary, an exciting ecological story is emerging, and one with clear management implications.
The South Taranaki Bight (STB) region of New Zealand is an important area for a population of blue whales which are unique to New Zealand. A wind-driven upwelling system brings cold, productive waters into the bight [1], which sustains high densities of krill [2], blue whale prey. The region is also frequented by busy shipping traffic, oil and gas drilling and extraction platforms as well as seismic survey effort for subsurface oil and gas reserves, and is the site of a recently-permitted seabed mine for iron sands (Fig. 1). However, a lack of knowledge on blue whale distribution and habitat use patterns has impeded effective management of these potential anthropogenic threats.
Figure 1. A blue whale surfaces in front of a floating production storage and offloading vessel servicing the oil rigs in the South Taranaki Bight. Photo by D. Barlow.
Three surveys were conducted in the STB region in the summer months of 2014, 2016, and 2017. During that time, we not only looked for blue whales, we also collected oceanographic data and hydroacoustic backscatter data to map and measure aspects of the krill in the region. These data streams will help us understand the functional, ecological relationships between the environment (oceanography), prey (krill), and predators (blue whales) in the ecosystem (Fig. 2). But in practice these data are costly and time-consuming to collect, while other data sources such as satellite imagery are readily accessible to managers at a variety of spatial and temporal scales. Therefore, another one of my aims is to link the data we collected in the field to satellite imagery, so that managers can have a practical tool to predict when and where the blue whales are most likely to be found in the region.
Figure 2. Data streams collected during surveys of the South Taranaki Bight Region in 2014, 2016, and 2017.
So what did I find? Here are the highlights from my preliminary analyses:
The majority of the patterns in blue whale distribution can be explained by the density, depth, and thickness of the krill patches.
Patterns in the krill are driven by oceanography.
Those same oceanographic parameters that drive the krill can be used to explain blue whale distribution.
There are tight relationships between the important oceanographic variables and satellite images of sea surface temperature.
Blue whale distribution can, to some degree, be explained using just satellite imagery.
We were able to identify a sea surface temperature range in the satellite imagery of approximately 18°C where the likelihood of finding a blue whale is the highest. Is this because blue whales really like 18° water? Well, more likely this relationship exists because the satellite imagery is reflective of the oceanography, and the oceanography drives patterns in the krill distribution, and the krill drives the distribution of blue whales (Fig. 3). We were able to make each of these functional linkages through our series of models, which is quite exciting.
Figure 3. The tiered modeling approach we took to investigate the ecological relationships between blue whales, krill, oceanography, and satellite imagery. Because of the ecological linkages we made, we are able to say that any relationship between whale distribution and satellite imagery most likely reflects a relationship between the blue whales and their prey.
That’s all well and good, but we were interested in testing these relationships to see if our identified habitat associations hold up even when we do not have field data (oceanographic, krill, and whale data). This past austral summer, we did not have a field season to collect data, but there was a large seismic airgun survey of the STB region. Seismic survey vessels are required to have trained marine mammal observers on board, and we were given access to the blue whale sightings data they recorded during the survey. In December, when the water was right around the preferred temperature identified by our models (18°C), the observers made 52 blue whale sightings (Fig. 4). In January and February, the waters warmed and only two sightings were made in each month. This is not only reassuring because it supports our model results, it also implies that there is the potential to balance industrial use of the area with protection of blue whale habitat, based on our understanding of the ecology. In January and February, very few blue whales were likely disturbed by the industrial activity in the STB, as conditions were not favorable for foraging at the location of the seismic survey. In contrast, the blue whales that were in the STB region in December may have experienced physiological consequences of sustained exposure to airgun noise since the conditions were favorable for foraging in the STB. In other words, the whales may have tolerated the noise exposure to gain access to good food, but this could have significant biological repercussions such as increased stress [3].
Figure 4. Monthly sea surface temperature (MODIS Aqua) overlaid with blue whale sightings from marine mammal observers aboard seismic survey vessel R/V Amazon Warrior. Black rectangles represent areas of seismic survey effort. Blue whale sighting location data were provided by RPS Energy Pty Ltd & Schlumberger, and Todd Energy.
In the first two weeks of July, we presented these latest findings to managers at the New Zealand Department of Conservation, the Minister of Conservation, the CEO and Policy Advisor of a major oil and gas conglomerate, NGOs, advocacy groups, and scientific colleagues. It was valuable to gather feedback from many different stakeholders, and satisfying to see such a clear interest in, and management application of, our work.
Dr. Leigh Torres and Dawn Barlow in front of Parliament in Wellington, New Zealand, following the presentation of their recent findings.
What’s next? We’re back in Oregon, and diving back into analysis. We intend to take the modeling work a step further to make the models predictive—for example, can we forecast where the blue whales will be based on the temperature, productivity, and winds two weeks prior? I am excited to see where these next steps lead!
References:
Shirtcliffe TGL, Moore MI, Cole AG, Viner AB, Baldwin R, Chapman B. 1990 Dynamics of the Cape Farewell upwelling plume, New Zealand. New Zeal. J. Mar. Freshw. Res.24, 555–568. (doi:10.1080/00288330.1990.9516446)
Bradford-Grieve JM, Murdoch RC, Chapman BE. 1993 Composition of macrozooplankton assemblages associated with the formation and decay of pulses within an upwelling plume in greater cook strait, New Zealand. New Zeal. J. Mar. Freshw. Res.27, 1–22. (doi:10.1080/00288330.1993.9516541)
Rolland RM, Parks SE, Hunt KE, Castellote M, Corkeron PJ, Nowacek DP, Wasser SK, Kraus SD. 2012 Evidence that ship noise increases stress in right whales. Proc. Biol. Sci.279, 2363–8. (doi:10.1098/rspb.2011.2429)



















































You must be logged in to post a comment.